Table of Contents |
The difference between prokaryotic cells and eukaryotic cells is often simply summarized with the statement that eukaryotic cells have a nucleus and prokaryotic cells do not. However, as you will learn in this lesson and in the lesson on eukaryotic cells, these cell types differ in many ways. It is important to understand these differences to understand how each type of cell functions. These differences are also very relevant in trying to develop medications that effectively treat bacterial infections while minimizing the risk to humans by targeting unique features of bacterial cells.
As discussed in previous lessons, prokaryotic cells are classified within two domains: Archaea and Bacteria. These two groups differ from each other in many ways. Although this lesson will focus on bacteria, it is important to remember that something that is true of bacterial cells may not be true of archaeal cells.
All cells contain cytoplasm (a gel-like substance composed of water and dissolved chemicals) and are bound by a phospholipid plasma membrane (also called a cell membrane or cytoplasmic membrane). All cells also contain chromosomes, which are complexes of DNA and proteins that help to organize the DNA. However, prokaryotic cells usually have a single, circular chromosome in a nucleoid region (not bound by a nucleus) and eukaryotic cells usually have multiple linear chromosomes inside a membrane-bound nucleus. All cells have ribosomes that make proteins, but the exact structure of the ribosomes differs by domain.
Prokaryotic and eukaryotic cells also divide differently. Cell division requires duplication of the chromosome or chromosomes. When one cell divides into two, the chromosomes must be properly distributed. You will learn more about this process and how it compares in prokaryotic and eukaryotic cells in the lesson on cell division and DNA replication.
IN CONTEXT
In addition to the examples mentioned above, there are other important ways that prokaryotic and eukaryotic cells differ. Eukaryotic cells are generally larger than prokaryotic cells, but there are some exceptions. Therefore, knowing the size of a cell is not sufficient to definitively classify it as a prokaryote or eukaryote. Prokaryotic cells may have membrane-bound structures, but they lack the complex system of membrane-bound compartments found in eukaryotic cells. Both eukaryotic and prokaryotic cells have a cytoskeleton, which consists of structures that help to give support and that are involved in movement. However, the cytoskeletal elements making up the cytoskeleton differ between them. There are also important differences in the ways that DNA is used to make proteins, in metabolism, in transport, and in other aspects of functioning.
The image below shows the major structures of a generic prokaryotic cell. These structures will be discussed in detail in this lesson. Note the oblong mass of DNA in the nucleoid region that is characteristic of a prokaryote.
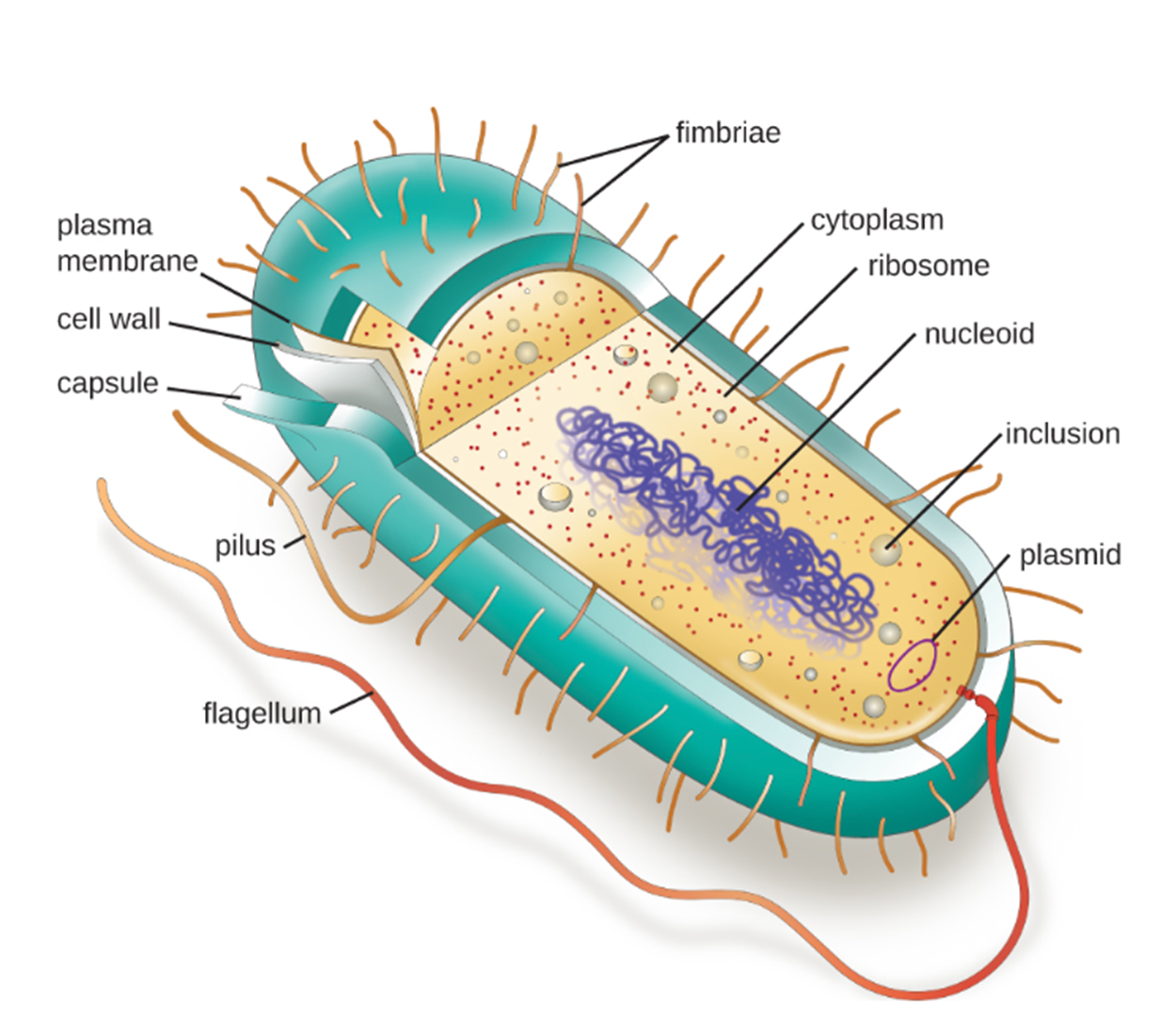
Individual cells of a particular prokaryotic organism are typically similar in shape (cell morphology). Although thousands of prokaryotic organisms have been identified, only a handful of cell morphologies are commonly seen microscopically. The table below summarizes common prokaryotic cell shapes.
| Common Prokaryotic Cell Shapes | |||
|---|---|---|---|
| Name | Description | Illustration | Image |
|
Coccus (pl. cocci) |
Round |

|
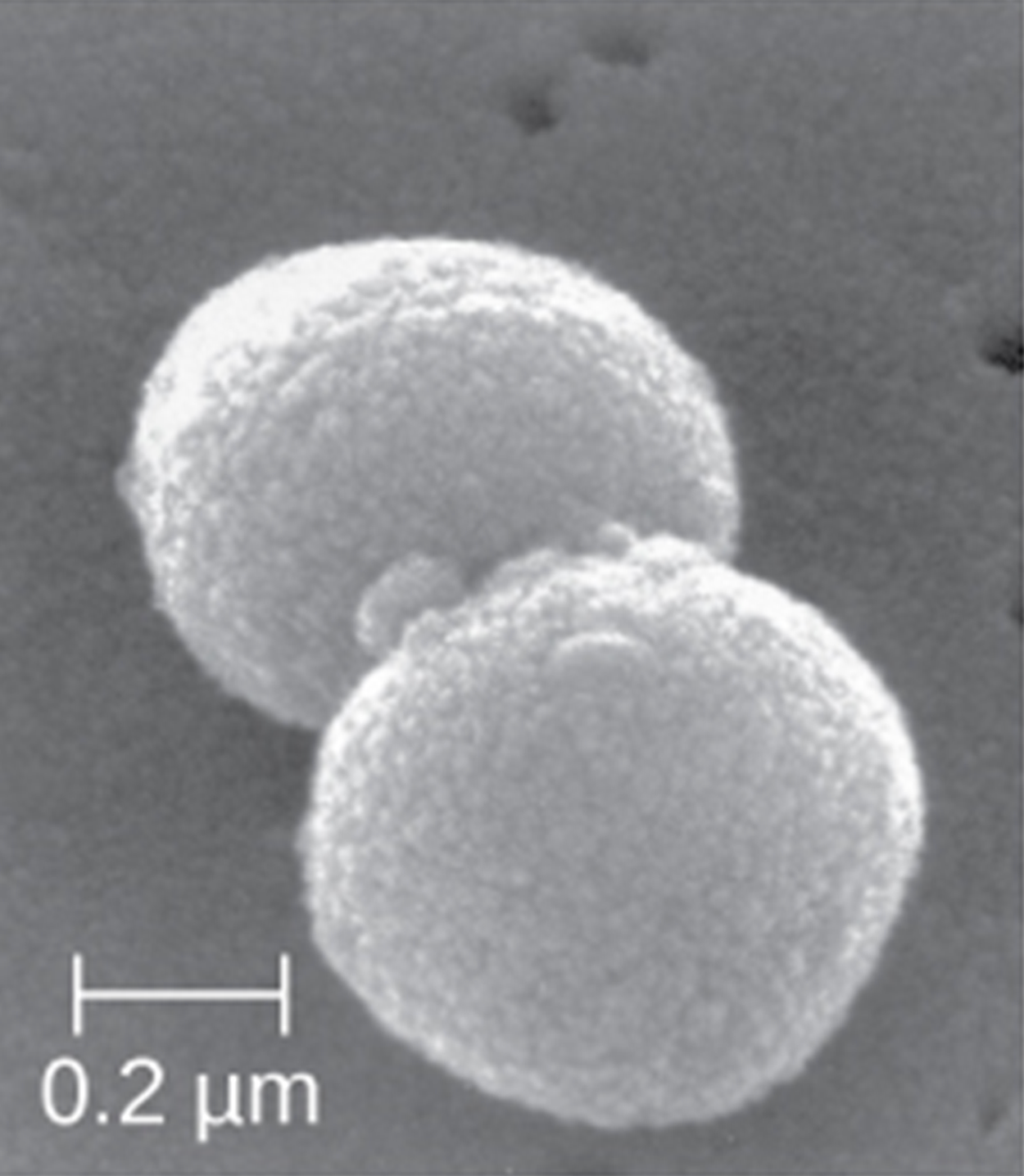
|
|
Bacillus (pl. bacilli) |
Rod |

|
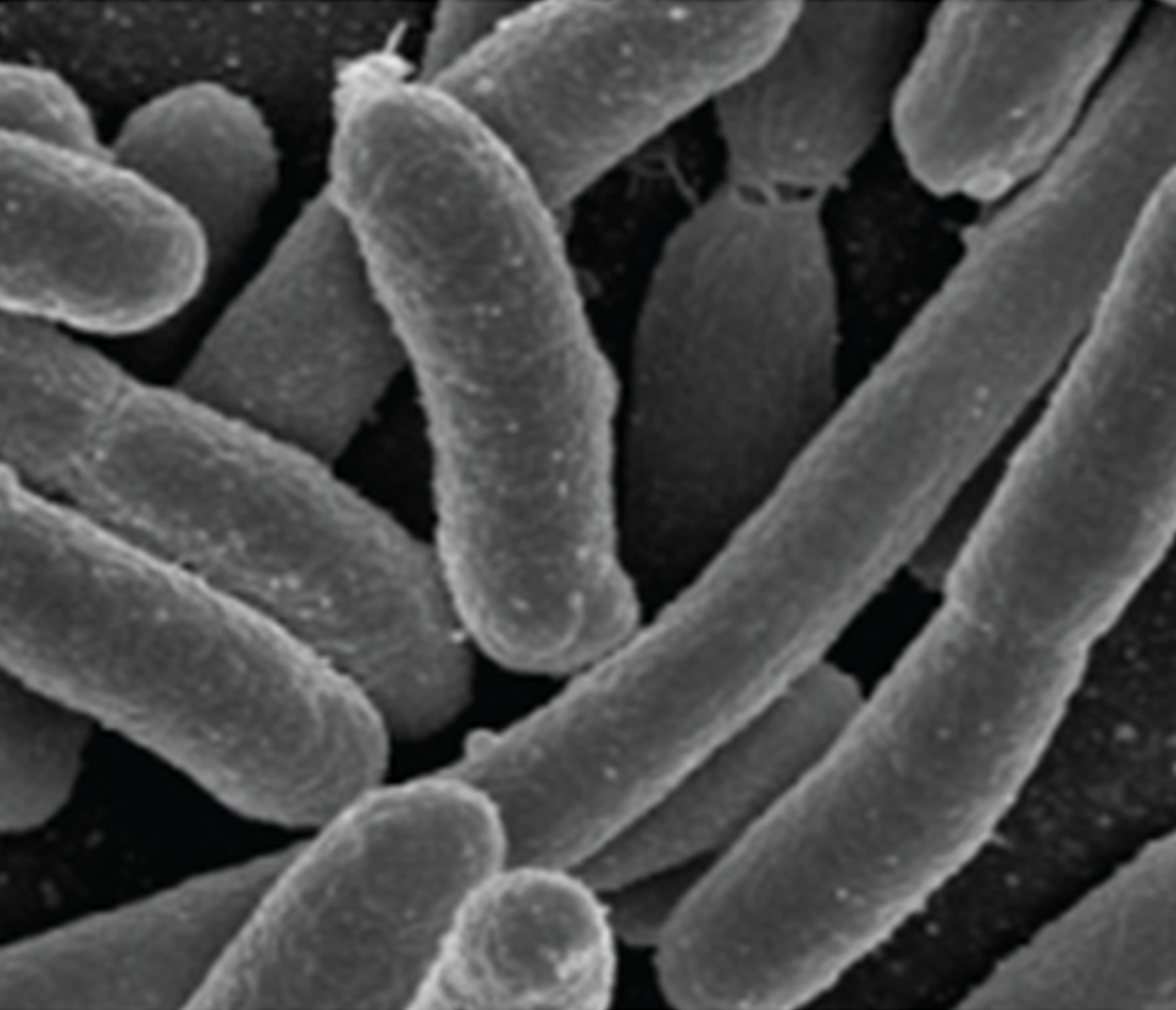
|
|
Vibrio (pl. vibrios) |
Curved rod |

|
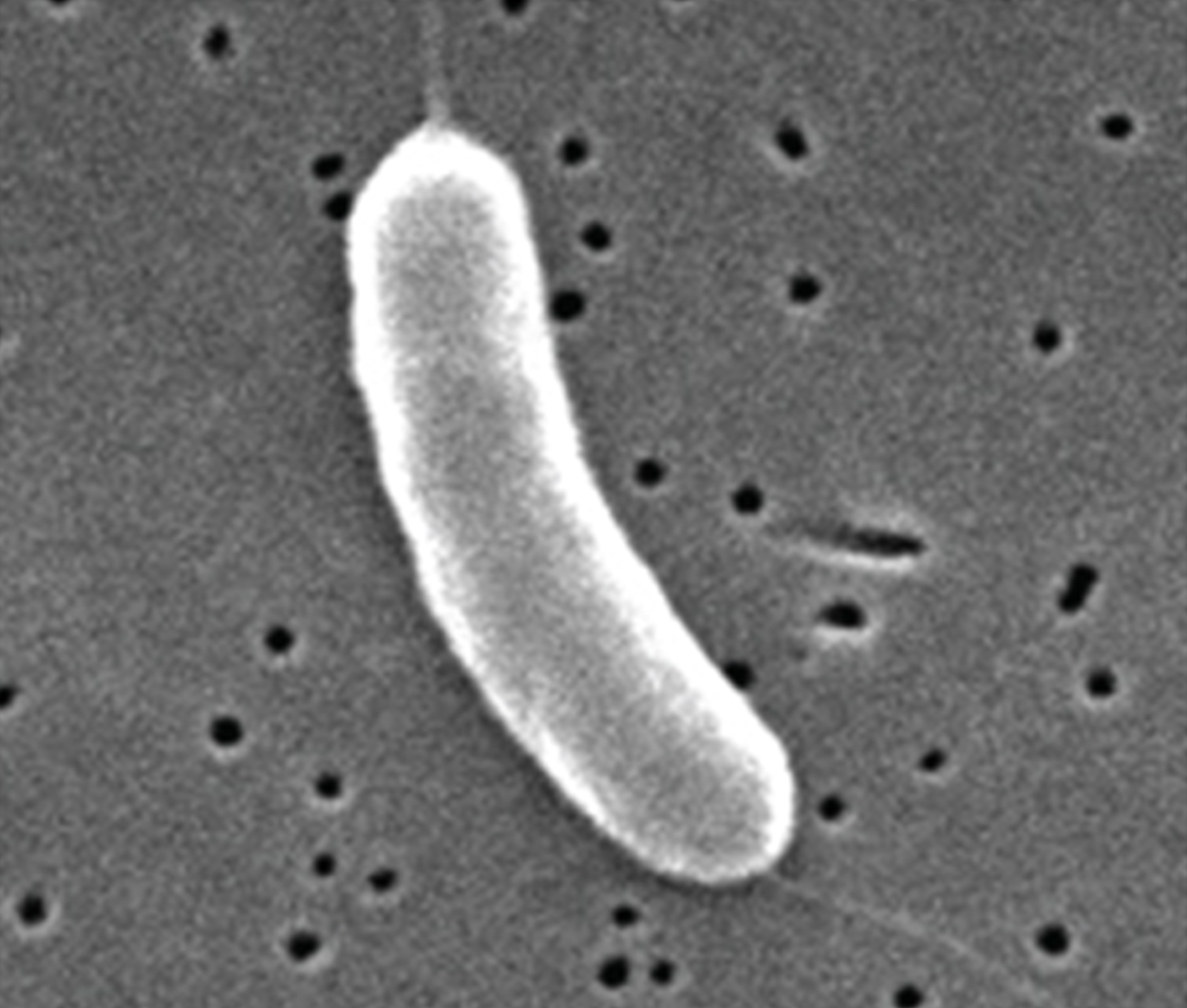
|
|
Coccobacillus (pl. coccobacilli) |
Short rod |

|
|
|
Spirillum (pl. spirilla) |
Spiral |

|
|
|
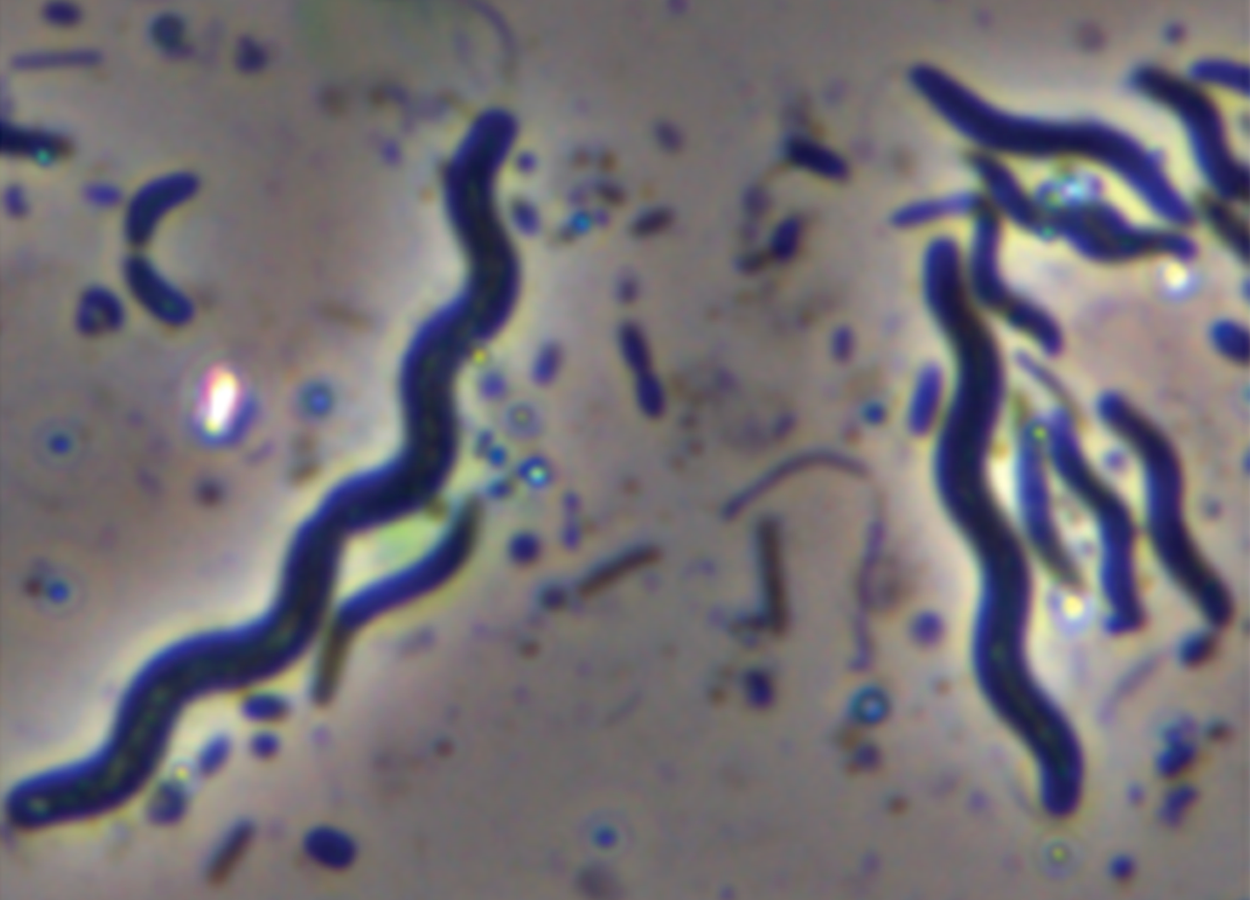
Spirochete (pl. spirochetes) |
Long, loose, helical spiral |
|
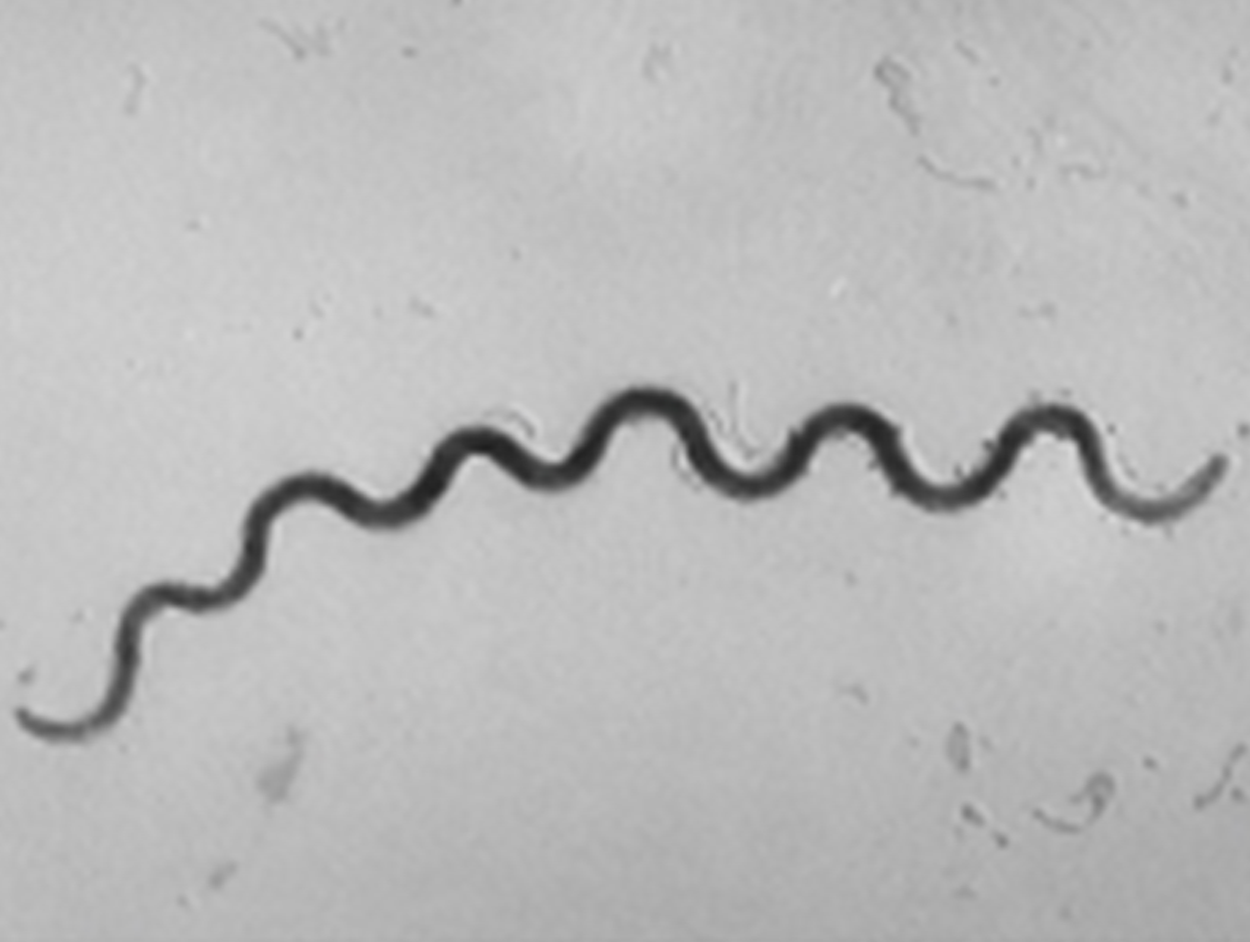
|
Some prokaryotic cells of the same species may group together in certain distinctive arrangements depending on the plane of cell division. Some common arrangements are shown in the table below.
| Common Prokaryotic Cell Arrangements | |||
|---|---|---|---|
| Name | Description | Illustration | |
|
Coccus (pl. cocci) |
Single coccus |

|
|
|
Diplococcus (pl. diplococci) |
Pair of two cocci |

|
|
|
Tetrad (pl. tetrads) |
Grouping of four cells arranged in a square |

|
|
|
Streptococcus (pl. streptococci) |
Chain of cocci |
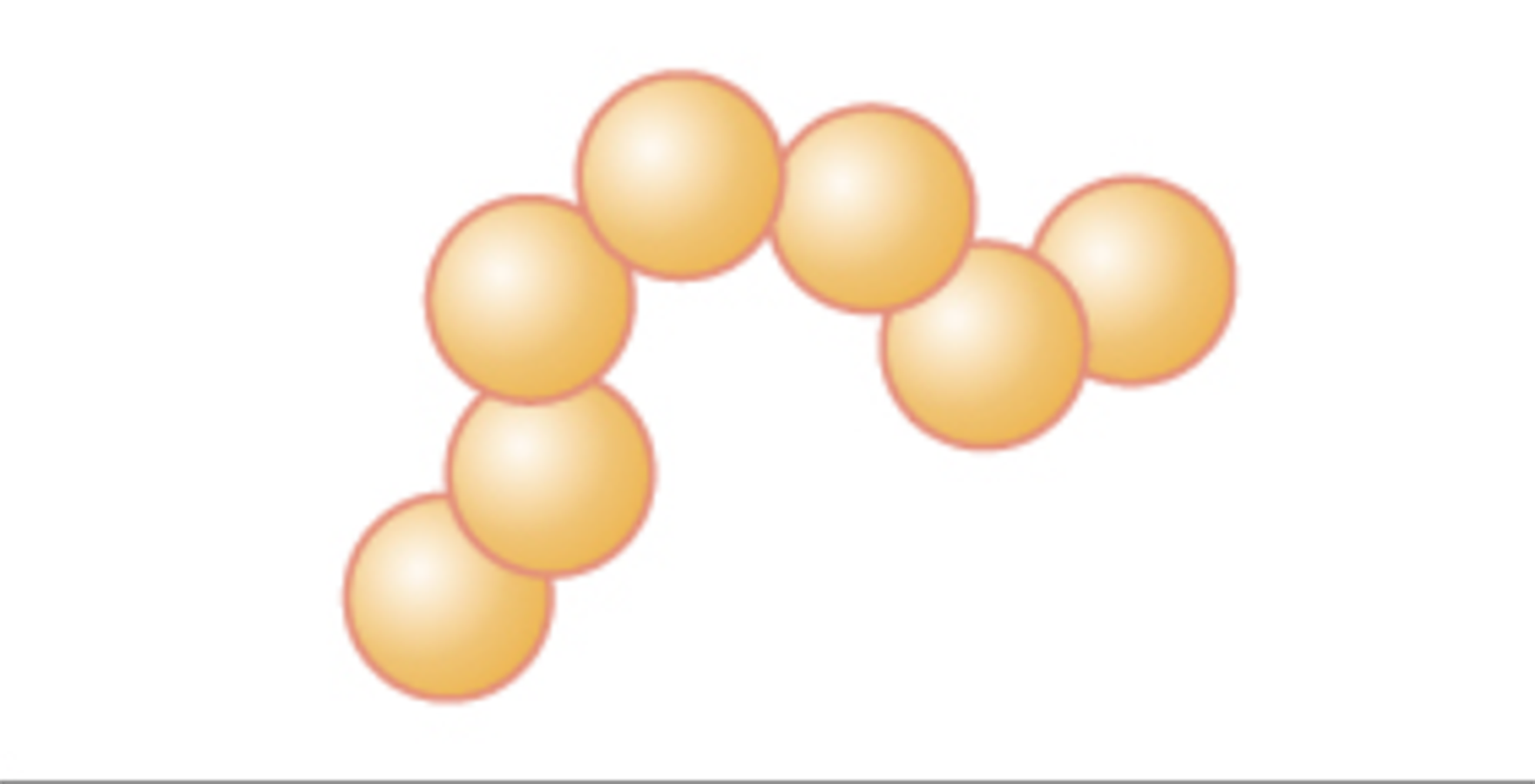
|
|
|
Staphylococcus (pl. staphylococci) |
Cluster of cocci |
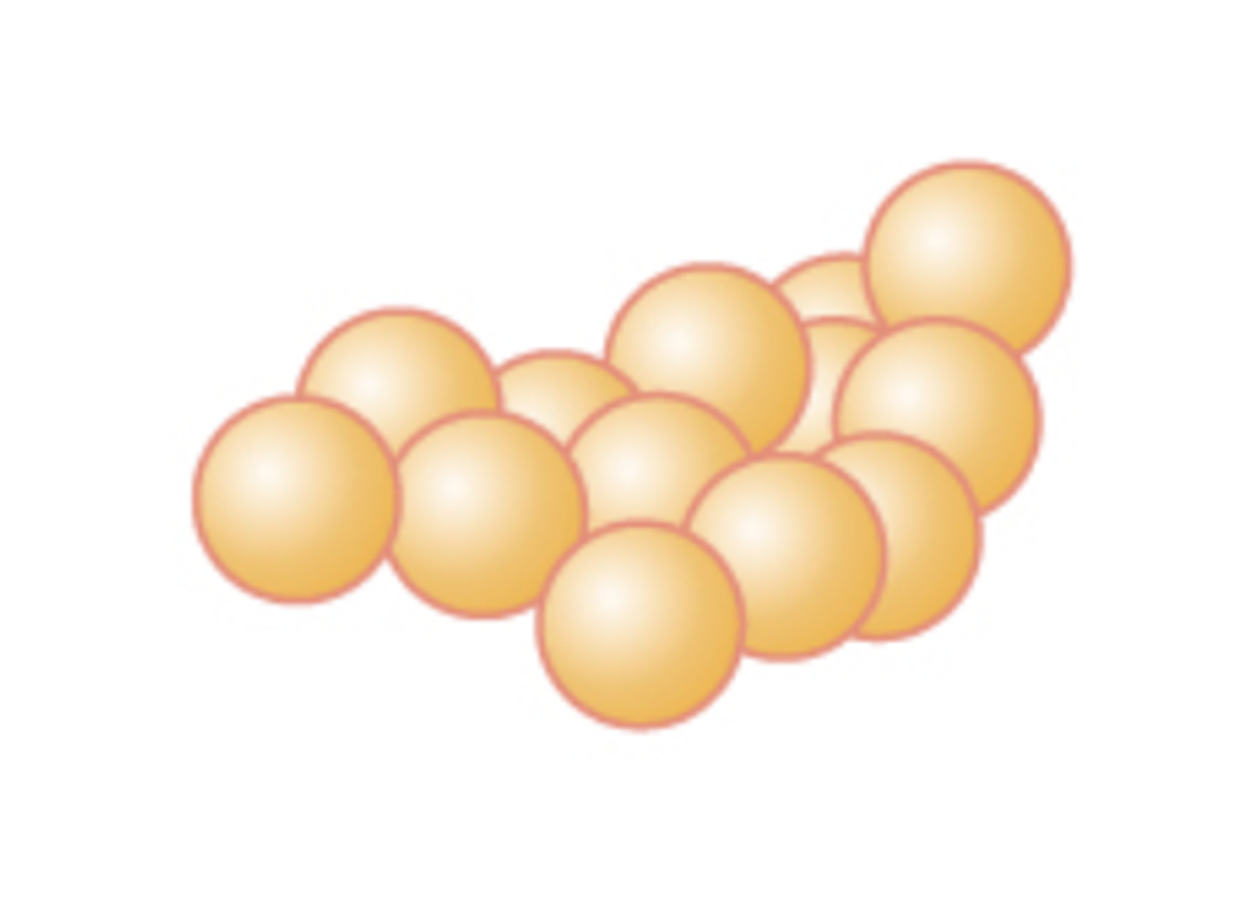
|
|
|
Bacillus (pl. bacilli) |
Single rod |

|
|
|
Streptobacillus (pl. streptobacilli) |
Chain of rods |
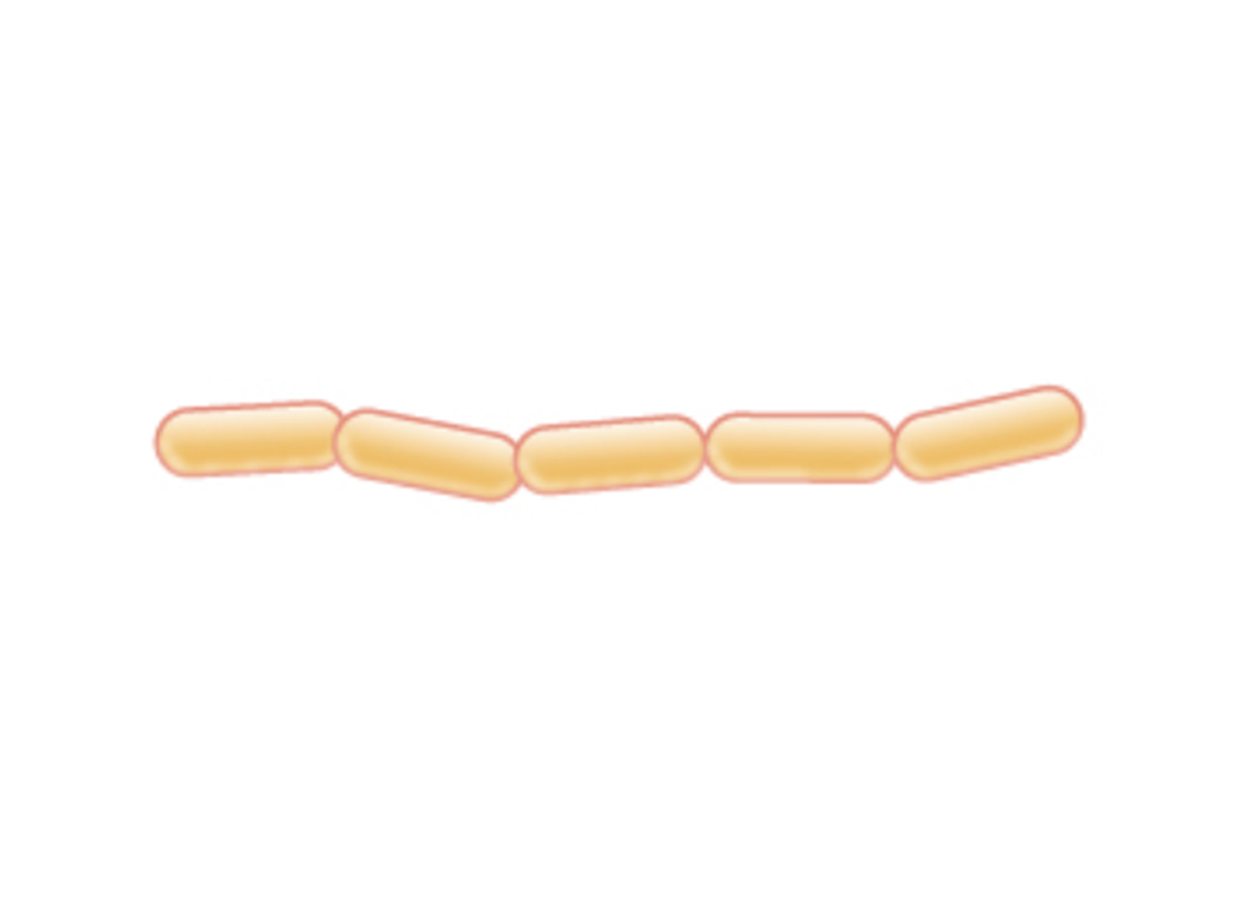
|
|
In most prokaryotic cells, morphology is maintained by the cell wall in combination with cytoskeletal elements. The cytoskeleton consists of structures that help to give support and that are involved in movement.
Most prokaryotic cells and some eukaryotic cells have a rigid cell wall located outside of the cell membrane. The cell wall provides structural strength and protection.
The cell wall helps to protect the cell from changes in osmotic pressure. Osmotic pressure is the pressure caused by water moving in response to differences in the concentrations of dissolved chemicals (solutes) on opposite sides of a semipermeable membrane (i.e., a membrane that allows some things to pass but not others). Osmosis is the movement of water across a semipermeable membrane as a result of differences in solute concentrations.
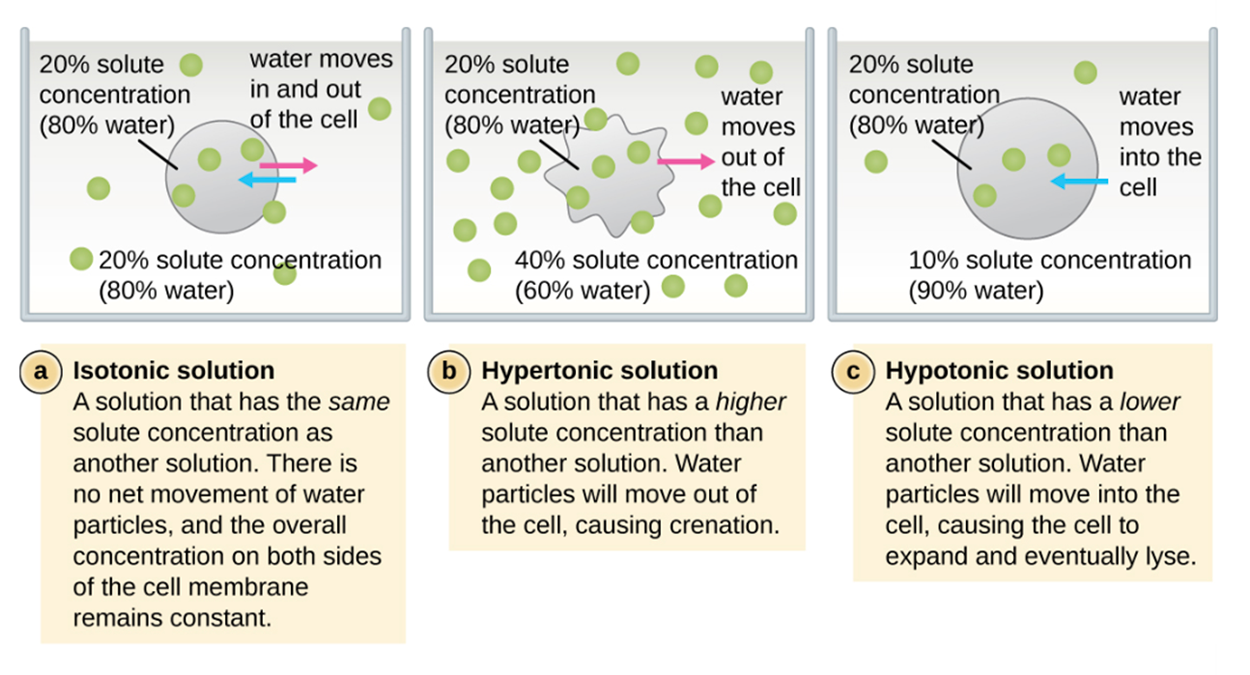
The external environment of a cell can be described as isotonic, hypertonic, or hypotonic and the type of environment determines whether there is a net gain or loss of water from the cell. Understanding the prefixes helps in understanding this process.
In an isotonic environment (“iso” means “equal” or “uniform”), the solute concentrations are similar inside and outside of the cell and there is no net movement of water. If the environment is hypertonic to the inside of the cell (“hyper” means “above”), then water flows out of the cell. Seawater is hypertonic to human cells and this is why humans can’t drink sea water when they need water. If the environment is hypotonic to the inside of the cell (“hypo” means “low”), then water flows into the cell and the cell can potentially lyse (burst) and die.
When a cell with no cell wall is placed in a hypertonic solution, it loses water and becomes shriveled. This is called crenation. When a cell with a cell wall is placed in a hypertonic solution, the plasma membrane shrinks away from the cell wall through a process called plasmolysis. In this case, the cell wall retains its shape.
Prokaryotic cells have a wide variety of internal structures. These include cytoskeletal elements, even though the cytoskeletal elements differ from those of eukaryotes. Particularly important structures include the nucleoid, plasmids, ribosomes, inclusions, and endospores.
The nucleoid is a region in a prokaryotic cell that contains prokaryotic DNA and DNA-associated proteins. Most prokaryotes have a single circular chromosome, although they may have additional small, circular pieces of DNA called plasmids found outside of the nucleoid region. In general, bacterial DNA interacts with nucleoid-associated proteins (NAPs) that assist in the organizing and packaging of the chromosome. These proteins have a similar function to histones, which are found in eukaryotes. In archaeans, the nucleoid is organized by either NAPs or histone-like DNA organizing proteins.
Prokaryotic cells may contain extrachromosomal DNA in addition to their main chromosome found in the nucleoid. This extrachromosomal DNA is found in plasmids, which are small, circular, double-stranded DNA molecules. Plasmids are more commonly found in bacteria, but have also been found in archaea and in eukaryotic organisms. Plasmids often carry genes that confer advantageous traits such as antibiotic resistance. You will learn more about plasmids in the lessons on genetics and biotechnology.
Ribosomes consist of ribosomal RNA (rRNA) and proteins. Prokaryotic ribosomes are found in the cytoplasm. They are called 70S ribosomes because they have a size of 70S, whereas eukaryotic cytoplasmic ribosomes have a size of 80S. (The S stands for Svedberg unit, a measure of sedimentation in an ultracentrifuge). Although they are the same size, bacterial and archaeal ribosomes have different proteins and rRNA molecules and archaeal ribosomes have important similarities to eukaryotic ribosomes.
Inclusions are structures used to store nutrients. There are many types of inclusions and they provide reservoirs of nutrients, in addition to other functions.
Some bacteria can form endospores, which are structures that help to protect the bacterial genome in a dormant state when conditions are unfavorable for growth. Endospores are resistant to extremes of temperature and radiation and can survive for extended periods before becoming metabolically active again (germination).
The process by which vegetative cells transform into endospores is called sporulation. It generally begins when nutrients become depleted or environmental conditions become otherwise unfavorable.
Structures that enclose the cytoplasm and internal structures of the cell are known collectively as the cell envelope. In prokaryotic cells, the structures of the cell envelope vary depending on the type of cell and organism. Most prokaryotic cells have a plasma membrane that exhibits selective permeability, allowing some materials through but not others.
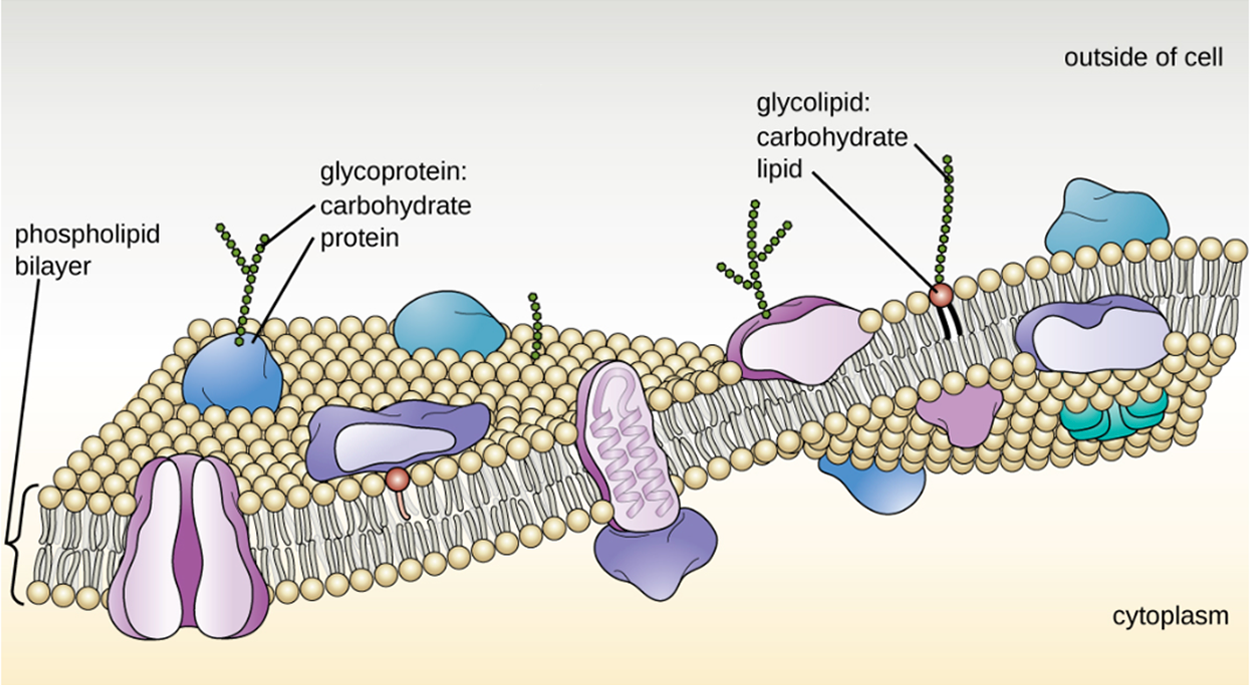
The fluid mosaic model is used to describe the structure of the cell membrane, which is shown in the image below. Most cell membranes consist of a phospholipid bilayer. There are proteins embedded in the membrane. The phospholipids and proteins can move to some extent within the membrane, although the amount of possible movement varies.
There are several especially important differences between archaeal and bacterial plasma membranes. First, archaeal membrane phospholipids are formed with ether linkages compared with ester linkages in bacteria. Secondly, archaeal phospholipids have branched chains whereas bacterial and eukaryotic phospholipids have straight chains. Some archaeans have a monolayer membrane, which may help to stabilize the membrane at high temperatures.
The phospholipids of the bilayer are arranged so that the outer and inner surfaces of the membrane are formed by hydrophilic (=“water loving”) phosphate heads and the inner part of the membrane is formed by hydrophobic (=“water hating”) lipid tails. Because “like dissolves like”, substances that dissolve well in water have difficulty passing through the lipid region. Small, uncharged molecules can pass through the membrane relatively easily. Small ions such as sodium ions cannot. However, lipids can pass more easily even if larger.
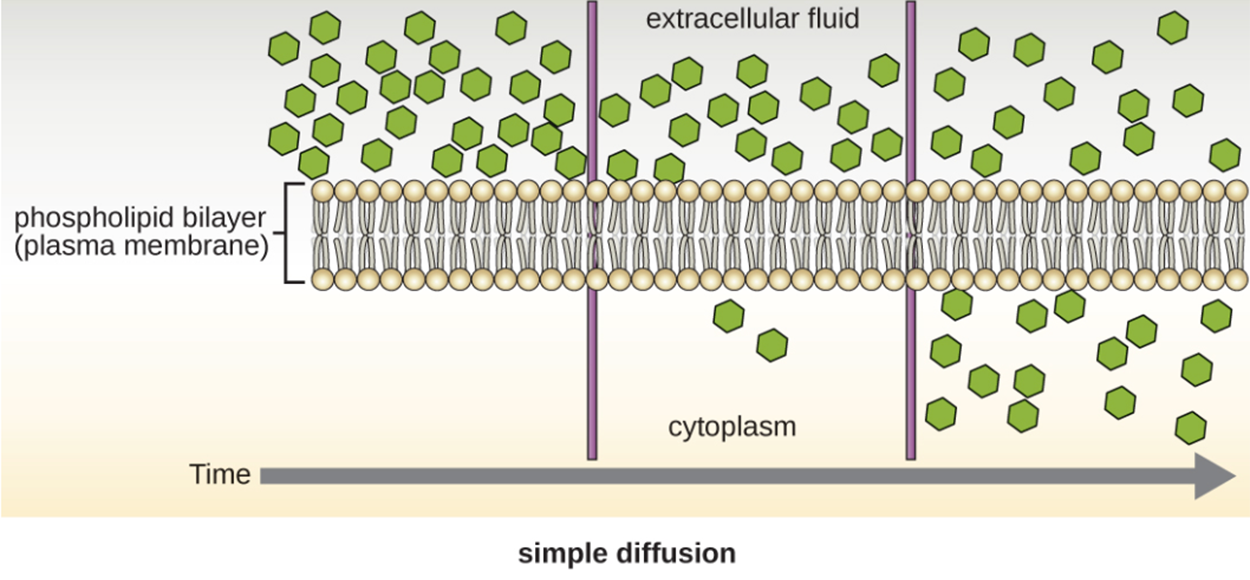
There are several ways by which substances cross the membrane. Some approaches occur spontaneously, but others require the presence of transport proteins with or without the use of energy.
Diffusion of molecules from an area of higher concentration to an area of lower concentration (i.e., down their concentration gradient) are transported by simple diffusion. This is called passive transport. Some small molecules, like carbon dioxide, can cross the membrane directly by simple diffusion. This is illustrated in the image below.
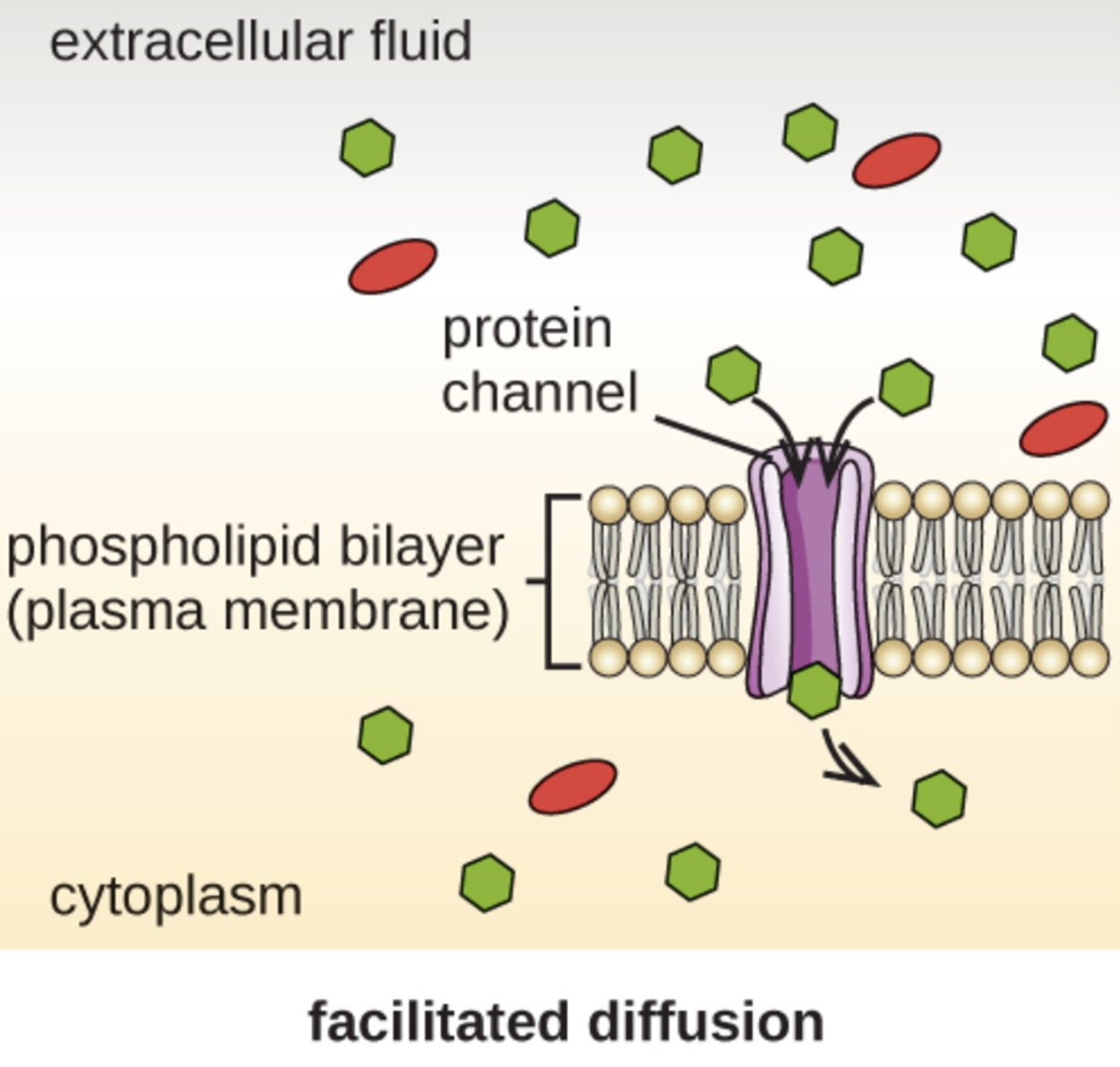
However, charged molecules and larger molecules may need the help of carrier or channel proteins. These structures help molecules across the membrane in a process called facilitated diffusion, which is illustrated in the image below.
Osmosis, discussed earlier in this lesson, is the diffusion of water across a semipermeable membrane and can occur through passive transport, but occurs more rapidly through facilitated transport by proteins called aquaporins. Note that a high concentration of water means a low concentration of solute and vice versa.
EXAMPLE
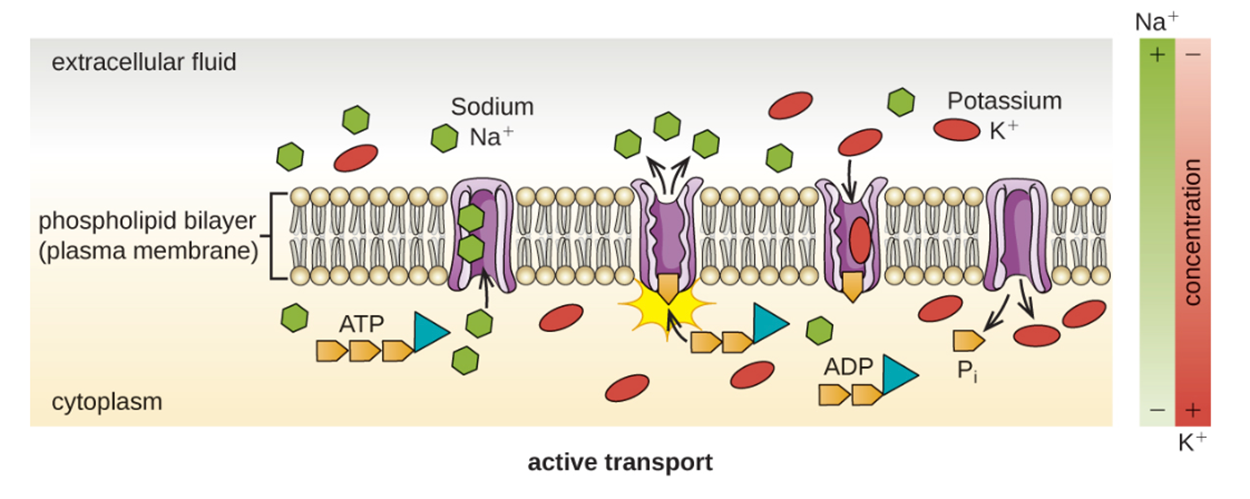
A hypertonic solution has a higher concentration of solute than the internal environment of a cell. This means that the concentration of water is lower outside the cell versus inside and there is a net movement of water outside of the cell.It is sometimes necessary to move molecules against a concentration gradient. In active transport, energy is used to move molecules from an area of low concentration to an area of high concentration. The proteins that perform active transport are often called pumps.
As already discussed, the cell wall is an important protective structure that helps cells maintain their shape despite osmotic pressure.
A major component of bacterial cell walls is peptidoglycan (murein), which is only found in bacteria. Peptidoglycan is targeted by some drugs, such as certain antibiotics, and some human immune cells recognize it.
Gram-positive and gram-negative cell walls differ. Gram-positive cell walls have a thicker peptidoglycan layer, generally containing teichoic acid carbohydrate chains that may help to stabilize them. Teichoic acid also plays a role in the ability of pathogenic gram-positive bacteria to bind to certain proteins on the surface of host cells, enhancing their ability to cause infection.
Bacteria of the family Mycobacteriaceae have an external layer of waxy mycolic acids in their cell walls as well, which is why they can be detected using acid-fast staining.
Gram-negative cells have a much thinner layer of peptidoglycan in their cell walls. Outside of the peptidoglycan layer, they have a periplasmic space and then a second lipid bilayer called the outer membrane. The outer membrane is attached to the peptidoglycan by the murein lipoprotein. The outer leaflet of the outer membrane contains the molecule lipopolysaccharide (LPS), which functions as an endotoxin in infections involving gram-negative bacteria. You will learn more about toxins in other lessons describing how pathogens cause disease.
Each LPS molecule is composed of Lipid A, a core polysaccharide, and an O side chain that is composed of sugar-like molecules. The composition of the O side chain varies between different species and strains of bacteria. Parts of the O side chain act as antigens (something to which the immune system may respond).
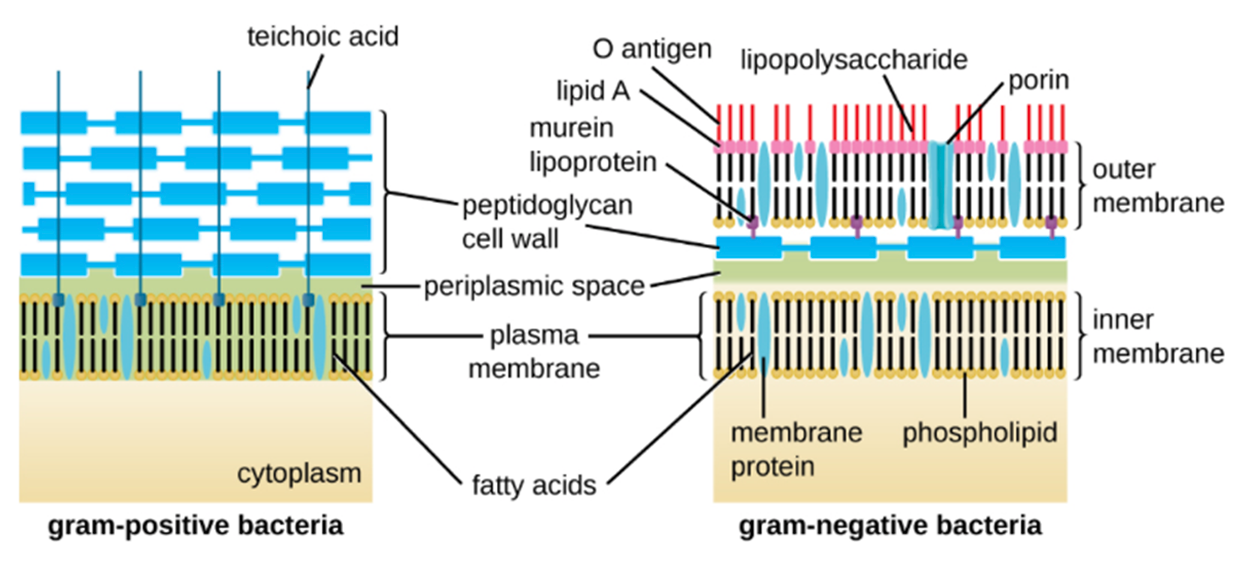
Archaeal cell wall structures have important differences from bacterial cell walls. They do not contain peptidoglycan. Instead, they often have a similar polymer called pseudopeptidoglycan (pseudomurein). Other archaea may have a layer of glycoproteins or polysaccharides that serves as the cell wall instead of pseudopeptidoglycan. A few species appear to lack cell walls.
Although most prokaryotic cells have cell walls, some may have additional cell envelope structures exterior to the cell wall. These structures include glycocalyces (singular=glycocalyx) and S-layers.
A glycocalyx is a carbohydrate outer covering. There are two major types: capsules and slime layers. A capsule is an organized layer located outside of the cell wall and usually composed of polysaccharides or proteins. They are not easily washed off. A slime layer is a less tightly organized layer that is only loosely attached to the cell wall and can be more easily washed off. Slime layers may be composed of polysaccharides, glycoproteins, or glycolipids.
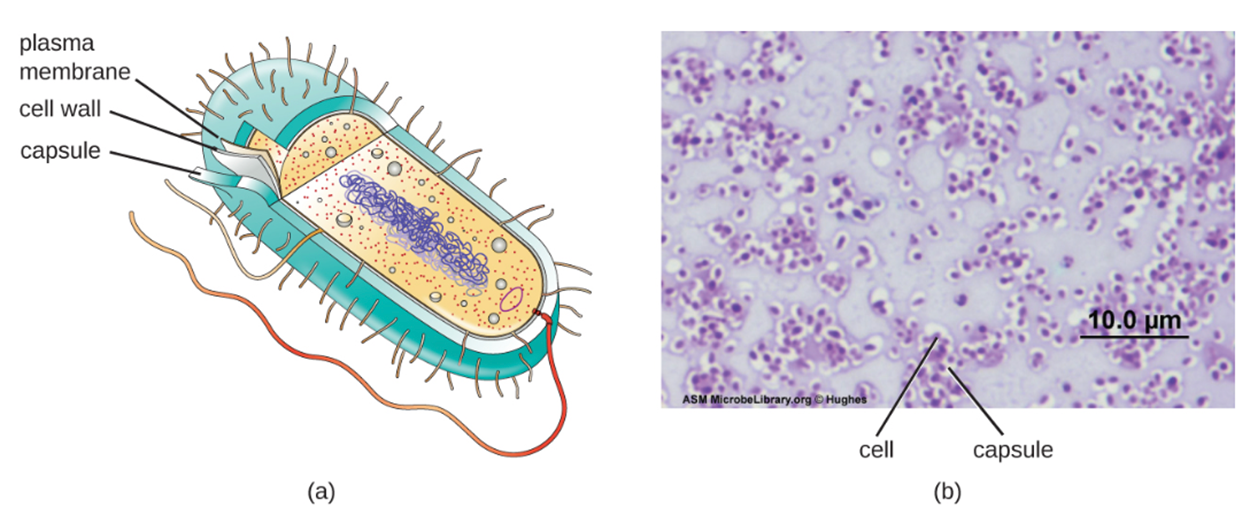
Glycocalyces allow cells to adhere to surfaces, aiding in the formation of biofilms (colonies of microbes that form in layers on surfaces). In nature, most bacteria live in mixed communities within biofilms. Biofilms provide protection from damage, desiccation, predation, and chemicals including antibiotics. Bacteria living in biofilms are more difficult to kill.
The ability to produce a capsule can contribute to a microbe’s pathogenicity (ability to cause disease) because the capsule can make it more difficult for immune cells to target the microbe.
An S-layer is another type of cell envelope structure. S-layers are composed of proteins and glycoproteins. In bacteria, S-layers are found outside of the cell wall. In some archaea, S-layers form the cell wall itself. The exact function of S-layers is uncertain, but they may provide protection from immune cells and osmotic pressure.
Many bacterial cells have protein appendages embedded within their cell envelopes that extend outward. These appendages can attach to other surfaces, transfer DNA, or allow movement. Filamentous appendages include fimbriae, pili, and flagella.
Fimbriae and pili are structurally similar and the terms are often used interchangeably. The term fimbriae commonly refers to short, bristle-like proteins projecting from the cell surface by the hundred. Fimbriae enable a cell to attach to surfaces and to other cells. For pathogenic bacteria, adherence to host cells is important for colonization, infectivity, and virulence. Adherence to surfaces is also important in biofilm formation. The term pili (singular: pilus) commonly refers to longer, less numerous protein appendages that aid in attachment to surfaces. A specific type of pilus, called the F pilus or sex pilus, is important for the transfer of DNA between bacterial cells. You will learn more about this process in the lesson on asexual reproduction and genetic diversity.
Flagella are structures used by cells to move in aqueous environments. Bacterial flagella act like propellers. They are stiff spiral filaments composed of flagellin protein subunits that extend outward from the cell and spin in solution. The basal body is the motor for the flagellum and is embedded in the plasma membrane. A hook region connects the basal body to the filament. Gram-positive and gram-negative bacteria have different basal body configurations and both have different flagella to eukaryotes.
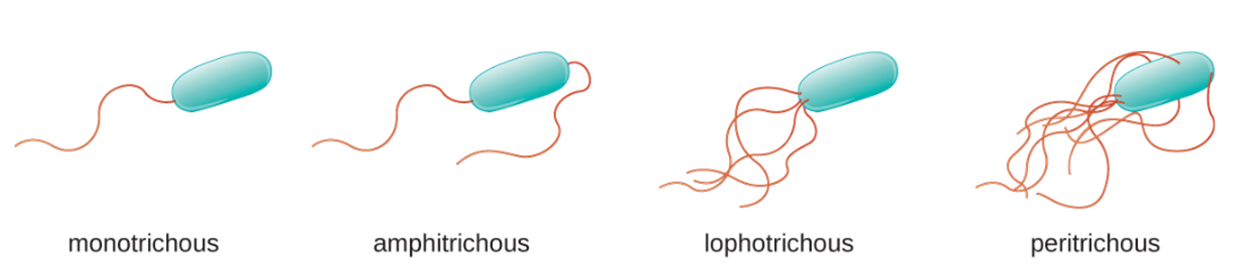
Flagella can be positioned in different arrangements as shown in the image below. A bacterium with a single flagellum, typically located at one end of the cell (polar), has a monotrichous flagellum. Cells with amphitrichous flagella have a flagellum or tufts of flagella at each end. Cells with lophotrichous flagella have a tuft at one end of the cell. Flagella that cover the entire surface of a bacterial cell are called peritrichous flagella.
Source: THIS TUTORIAL HAS BEEN ADAPTED FROM OPENSTAX "MICROBIOLOGY." ACCESS FOR FREE AT openstax.org/details/books/microbiology. LICENSE: CC ATTRIBUTION 4.0 INTERNATIONAL. Accessed by August 2022.
REFERENCES
Merriam-Webster. (n.d.). Is. In Merriam-Webster.com dictionary. Retrieved August 10, 2022, from www.merriam-webster.com/dictionary/is.
Merriam-Webster. (n.d.). Hyper. In Merriam-Webster.com dictionary. Retrieved August 10, 2022, from www.merriam-webster.com/dictionary/hyper.
Merriam-Webster. (n.d.). Hypo. In Merriam-Webster.com dictionary. Retrieved August 10, 2022, from www.merriam-webster.com/dictionary/hypo.
Parker, N., Schneegurt, M., Thi Tu, A.-H., Lister, P., & Forster, B. (2016). Microbiology. OpenStax. Access for free at openstax.org/books/microbiology/pages/1-introduction.