Table of Contents |
The immune system is a collection of barriers, cells, and soluble proteins that interact and communicate with each other in extraordinarily complex ways. The modern model of immune function is organized into three phases based on the timing of their effects. The three temporal phases consist of the following:

| Term | Pronunciation | Audio File |
|---|---|---|
| Lymphocytes | lym·pho·cytes |
|
As stated above, lymphocytes are the primary cells of adaptive immune responses. The two basic types of lymphocytes, B cells and T cells, are morphologically identical, with a large central nucleus surrounded by a thin layer of cytoplasm. They are distinguished from each other by their surface protein markers as well as by the molecules they secrete. While B cells mature in red bone marrow and T cells mature in the thymus, they both initially develop from bone marrow. T cells migrate from bone marrow to the thymus gland where they further mature. B cells and T cells are found in many parts of the body, circulating in the bloodstream and lymph, and residing in secondary lymphoid organs, including the spleen and lymph nodes, which will be described in a future lesson. The human body contains approximately 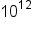 lymphocytes. The types of lymphocytes and their primary functions are shown in the table below.
lymphocytes. The types of lymphocytes and their primary functions are shown in the table below.
| Type of lymphocyte | Primary function |
|---|---|
| B lymphocyte | Generates diverse antibodies |
| Plasma cell | Secretes antibodies |
| T lymphocyte | Secretes chemical messengers |
| Natural killer (NK) cell | Destroys virally infected cells |
B cells are immune cells that function primarily by producing antibodies. An antibody is any of the group of proteins that specifically binds to pathogen-associated molecules known as antigens. An antigen is a chemical structure on the surface of a pathogen that binds to T or B lymphocyte antigen receptors. Once activated by binding to antigen, B cells differentiate into cells that secrete a soluble form of their surface antibodies. These activated B cells are known as plasma cells, which secrete antibodies.
T cells, on the other hand, do not secrete antibodies, but rather perform a variety of functions in the adaptive immune response. Different T cell types have the ability to either secrete soluble factors that communicate with other cells of the adaptive immune response or destroy cells infected with intracellular pathogens. The roles of T and B lymphocytes in the adaptive immune response will be discussed further in this lesson.
A fourth important lymphocyte is the natural killer cell, which participates in the innate immune response. A natural killer (NK) cell is a circulating blood cell that contains cytotoxic (cell-killing) granules in its extensive cytoplasm. It shares this mechanism with the cytotoxic T cells of the adaptive immune response. NK cells are among the body’s first lines of defense against viruses and certain types of cancer.
The process of eliminating T cells that might attack the cells of one’s own body is referred to as T cell tolerance. While thymocytes are in the cortex of the thymus, they are referred to as “double negatives,” meaning that they do not bear the CD4 or CD8 molecules that you can use to follow their pathways of differentiation. In the cortex of the thymus, they are exposed to cortical epithelial cells. In a process known as positive selection, double-negative thymocytes bind to the MHC molecules they observe on the thymic epithelia, and the MHC molecules of “self” are selected. This mechanism kills many thymocytes during T cell differentiation. In fact, only 2% of the thymocytes that enter the thymus leave it as mature, functional T cells.

Later, the cells become double positives that express both CD4 and CD8 markers and move from the cortex to the junction between the cortex and medulla. It is here that negative selection takes place. In negative selection, self-antigens are brought into the thymus from other parts of the body by professional antigen-presenting cells. The T cells that bind to these self-antigens are selected negatively and are killed by apoptosis, which is cell death induced by a cell’s own internal mechanisms. In summary, the only T cells left are those that can bind to MHC molecules of the body with foreign antigens presented on their binding clefts, preventing an attack on one’s own body tissues, at least under normal circumstances. Tolerance can be broken, however, by the development of an autoimmune response, to be discussed later in this chapter.
The cells that leave the thymus become single positives, expressing either CD4 or CD8, but not both (see the figure above). The CD4⁺ T cells will bind to MHC class II molecules and the CD8⁺ cells will bind to MHC class I molecules. The discussion that follows explains the functions of these molecules and how they can be used to differentiate between the different T cell functional types.
B cells differentiate in the bone marrow. During the process of maturation, up to 100 trillion different clones of B cells are generated, which is similar to the diversity of antigen receptors seen in T cells.
B cell differentiation and the development of tolerance are not quite as well understood as it is in T cells. Central tolerance is the destruction or inactivation of B cells that recognize self-antigens in the bone marrow, and its role is critical and well-established. In the process of clonal deletion, immature B cells that bind strongly to self-antigens expressed on tissues are signaled to induce their own destruction by apoptosis, removing them from the population. In the process of clonal anergy, however, B cells exposed to soluble antigens in the bone marrow are not physically deleted but become unable to function.
Another mechanism called peripheral tolerance is a direct result of T cell tolerance. In peripheral tolerance, functional, mature B cells leave the bone marrow but have yet to be exposed to self-antigen. Most protein antigens require signals from helper T (Th2) cells to proceed to make antibodies. When a B cell binds to a self-antigen but receives no signals from a nearby Th2 cell to produce antibodies, the cell is signaled to undergo apoptosis and is destroyed. This is yet another example of the control that T cells have over the adaptive immune response.
After B cells are activated by binding to antigen, they differentiate into plasma cells. Plasma cells often leave the secondary lymphoid organs, where the response is generated, and migrate back to the bone marrow, where the whole differentiation process started. After secreting antibodies for a specific period, they die, as most of their energy is devoted to making antibodies and not to maintaining themselves. Thus, plasma cells are said to be terminally differentiated.
The final B cell of interest is the memory B cell, which results from the clonal expansion of an activated B cell. Memory B cells function in a way similar to memory T cells. They lead to a stronger and faster secondary response compared with the primary response.
| Clonal Anergy | c·lon·al an·er·gy |
|
Source: THIS TUTORIAL HAS BEEN ADAPTED FROM OPENSTAX "ANATOMY AND PHYSIOLOGY 2E" ACCESS FOR FREE AT OPENSTAX.ORG/DETAILS/BOOKS/ANATOMY-AND-PHYSIOLOGY-2E. LICENSE: CREATIVE COMMONS ATTRIBUTION 4.0 INTERNATIONAL